


O pesquisador escocês Robert Brown (1773- 1858) é
considerado o descobridor do núcleo celular. Embora muitos citologistas
anteriores a ele já tivessem observados núcleos, não haviam compreendido
a enorme importância dessas estruturas para a vida das células. O
grande mérito de Brown foi justamente reconhecer o núcleo como
componente fundamental das células. O nome que ele escolheu expressa
essa convicção: a palavra “núcleo” vem do grego nux, que significa semente. Brown imaginou que o núcleo fosse a semente da célula, por analogia aos frutos.

Hoje, sabemos que o núcleo é o centro de controle
das atividades celulares e o “arquivo” das informações hereditárias,
que a célula transmite às suas filhas ao se reproduzir.
Células eucariontes e procariontes
A membrana celular presente nas células
eucariontes, mas ausente nas procariontes. Na célula eucarionte, o
material hereditário está separado do citoplasma por uma membrana – a
carioteca – enquanto na célula procarionte o material hereditário se
encontra mergulhado diretamente no líquido citoplasmático.
Os componentes do núcleo
O núcleo das célula que não estão em processo de divisão apresenta um limite bem definido, devido à presença da carioteca ou membrana nuclear, visível apenas ao microscópio eletrônico.
A maior parte do volume nuclear é ocupada por uma massa filamentosa denominada cromatina. Existem ainda um ou mais corpos densos (nucléolos) e um líquido viscoso (cariolinfa ou nucleoplasma).
A carioteca
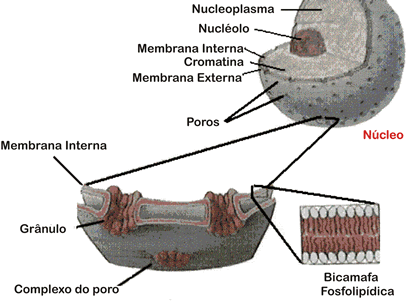 A carioteca (do grego karyon, núcleo e theke,
invólucro, caixa) é um envoltório formado por duas membranas
lipoprotéicas cuja organização molecular é semelhante as demais
membranas celulares. Entre essas duas membranas existe um estreito
espaço, chamado cavidade perinuclear.
A carioteca (do grego karyon, núcleo e theke,
invólucro, caixa) é um envoltório formado por duas membranas
lipoprotéicas cuja organização molecular é semelhante as demais
membranas celulares. Entre essas duas membranas existe um estreito
espaço, chamado cavidade perinuclear.
A face externa da carioteca, em algumas partes,
se comunica com o retículo endoplasmático e, muitas vezes, apresenta
ribossomos aderidos à sua superfície. Neste caso, o espaço entre as duas
membranas nucleares é uma continuação do espaço interno do retículo
endoplasmático.
A carioteca é perfurada por milhares de poros,
através das quais determinadas substâncias entram e saem do núcleo. Os
poros nucleares são mais do que simples aberturas. Em cada poro existe
uma complexa estrutura protéica que funciona como uma válvula,
abrindo-se para dar passagem a determinadas moléculas e fechando-se em
seguida. Dessa forma, a carioteca pode controlar a entrada e a saída de
substâncias.

A face interna da carioteca encontra-se a lâmina
nuclear, uma rede de proteínas que lhe dá sustentação. A lâmina nuclear
participa da fragmentação e da reconstituição da carioteca, fenômenos
que ocorrem durante a divisão celular.
A cromatina
A cromatina (do grego chromatos, cor) é
um conjunto de fios, cada um deles formado por uma longa molécula de DNA
associada a moléculas de histonas, um tipo especial de proteína. Esses
fios são os cromossomos.
Quando se observam núcleos corados ao microscópio
óptico, nota-se que certas regiões da cromatina se coram mais
intensamente do que outras. Os antigos citologistas já haviam observados
esse fato e imaginado, acertadamente, que as regiões mais coradas
correspondiam a porções dos cromossomos mais enroladas, ou mais
condensadas, do que outras.
Para assinalar diferenças entre os tipos de cromatina, foi criado o termo heterocromatina (do grego heteros,
diferente), que se refere à cromatina mais densamente enrolada. O
restante do material cromossômico, de consistência mais frouxa, foi
denominado eucromatina (do grego eu, verdadeiro).

Diferentes níveis de condensação do DNA. (1) Cadeia simples de DNA . (2) Filamento de cromatina (DNA com histonas). (3) Cromatina condensada em interfase com centrómeros. (4) Cromatina condensada em profase. (Existem agora duas cópias da molécula de DNA) (5) Cromossoma em metafase
Os nucléolos
Na fase que a célula eucariótica não se encontra
em divisão é possível visualizas vários nucléolos, associados a algumas
regiões específicas da cromatina. Cada nucléolo é um corpúsculo
esférico, não membranoso, de aspecto esponjoso quando visto ao
microscópio eletrônico, rico em RNA ribossômico (a sigla RNA provém do inglês RiboNucleic Acid).
Este RNA é um ácido nucléico produzido a partir o DNA das regiões
específicas da cromatina e se constituirá um dos principais componentes
dos ribossomos presentes no citoplasma.
É importante perceber que ao ocorrer a
espiralação cromossômica os nucléolos vão desaparecendo lentamente. Isso
acontece durante os eventos que caracterizam a divisão celular. O
reaparecimento dos nucléolos ocorre com a desespiralação dos
cromossomos, no final da divisão do núcleo.
Cromossomos da célula interfásica
O período de vida da célula em que ela
não está em processo de divisão é denominado interfase. A cromatina da
célula interfásica, como já foi mencionada, é uma massa de filamentos
chamados de cromossomos. Se pudéssemos separar, um por um, os
cromossomos de uma célula interfásica humana, obteríamos 46 filamentos,
logos e finos. Colocado em linha, os cromossomos humanos formariam um
fio de 5 cm de comprimento, invisível ao microscópio óptico, uma vez que
sua espessura não ultrapassa 30 nm.

Constituição química e arquitetura dos cromossomos
Descobrir a natureza química dos cromossomos foi
uma árdua tarefa que mobilizou centenas de cientistas e muitos anos de
trabalho. O primeiro constituinte cromossômico a ser identificado foi o
ácido desoxirribonucléico, o DNA.
Em 1924, o pesquisador alemão Robert J. Feugen
desenvolveu uma técnica especial de coloração que permitiu demonstrar
que o DNA é um dos principais componentes dos cromossomos. Alguns anos
mais tarde, descobriu-se que a cromatina também é rica em proteínas
denominadas histonas.
Cromossomos da célula em divisão
Quando a célula vai se dividir, o núcleo e os
cromossomos passam por grandes modificações. Os preparativos para a
divisão celular têm inicio com a condensação dos cromossomos, que
começam a se enrolar sobre si mesmos, tornando-se progressivamente mais
curtos e grossos, até assumirem o aspecto de bastões compactos.
Constrições cromossômicas
Durante a condensação cromossômica, as regiões
eucromáticas se enrolam mais frouxamente do que as heterocromáticas, que
estão condensadas mesmo durante a interfase. No cromossomo condensado,
as heterocromatinas, devido a esse alto grau de empacotamento, aparecem
como regiões “estranguladas” do bastão cromossômico, chamadas
constrições.

Na célula que está em processo de divisão, cada
cromossomo condensado aparece como um par de bastões unidos em um
determinado ponto, o centrômero. Essas duas “metades” cromossômicas,
denominadas cromátides-irmãs são idênticas e surgem da duplicação do
filamento cromossômico original, que ocorre na interfase, pouco antes de
a divisão celular se iniciar.
Durante o processo de divisão celular, as
cromátides-irmãs se separam: cada cromátide migra para uma das
células-filhas que se formam.
O centrômero fica localizado em uma região
heterocromática, portanto em uma constrição que contém o centrômero é
chamada constrição primária, e todas as outras que porventura existam
são chamadas constrições secundárias.

As partes de um cromossomo separadas pelo
centrômero são chamadas braços cromossômicos. A relação de tamanho entre
os braços cromossômicos, determinada pela posição do centrômero,
permite classificar os cromossomos em quatro tipos:
- metacêntrico: possuem o centrômero no meio, formando dois braços de mesmo tamanho;
- submetacêntricos: possuem o centrômero um pouco deslocado da região mediana, formando dois braços de tamanhos desiguais;
- acrocêntricos: possuem o centrômero bem próximo a uma das extremidades, formando um braço grande e outro muito pequeno;
- telocêntricos: possuem o centrômero em um das extremidades, tendo apenas um braço.

Cromossomos e genes
As moléculas de DNA dos cromossomos contêm
“receitas” para a fabricação de todas as proteínas da célula. Cada
“receita” é um gene.
Portanto, o gene é uma seqüência de nucleotídeos do DNA que pode ser transcrita em uma versão de RNA e conseqüentemente traduzida em uma proteína.

Conceito de genoma
Um cromossomo é comparável a um livro de receita
de proteínas, e o núcleo de uma célula humana é comparável a uma
biblioteca, constituída por 46 volumes, que contêm o receituário
completo de todas as proteínas do indivíduo. O conjunto completo de
genes de uma espécie, com as informações para a fabricação dos milhares
de tipos de proteínas necessários à vida, é denominado genoma.
Atualmente, graças a modernas técnicas de identificação dos genes, os
cientistas mapearam o genoma humano através do Projeto Genoma Humano.
Projeto Genoma Humano
O Projeto Genoma Humano (PGH)
teve por objetivo o mapeamento do genoma humano, e a identificação de
todos os nucleotídeos que o compõem. Consistiu num esforço mundial para
se decifrar o genoma. Após a iniciativa do National Institutes of Health
(NIH) dos Estados Unidos, centenas de laboratórios de todo o mundo se
uniram à tarefa de seqüenciar, um a um, os genes que codificam as
proteínas do corpo humano e também aquelas seqüências de DNA que não são
genes. Laboratórios de países em desenvolvimento também participaram do
empreendimento com o objetivo de formar mão-de-obra qualificada em
genômica.
 |
Para o seqüenciamento de um gene, é necessário que ele seja antes amplificado numa reação em cadeia da polimerase,
e então clonado em bactérias. Após a obtenção de quantidade suficiente
de DNA, executa-se uma nova reação em cadeia (PCR), desta vez utilizando
didesoxirribonucleotídeos marcados com fluoróforos para a determinação da seqüência.
O projeto foi fundado em 1990, com um
financiamento de 3 milhões de dólares do Departamento de Energia dos
Estados Unidos e dos Institutos Nacionais de Saúde dos Estados Unidos, e
tinha um prazo previsto de 15 anos.
|
Devido à grande cooperação da comunidade
científica internacional, associada aos avanços no campo da
bioinformática e das tecnologias de informação, um primeiro esboço do
genoma foi anunciado em 26 de Junho de 2000, dois anos antes do
previsto.
Em 14 de Abril de 2003, um comunicado de
imprensa conjunto anunciou que o projeto foi concluído com sucesso, com o
seqüenciamento de 99% do genoma humano, com uma precisão de 99,99%.
Os trabalhos do projeto foram dados como
concluídos em 2003. Com a tecnologia da época, estimou-se que todos os
genes (em torno de 25.000) haviam sido seqüenciados. Deve-se lembrar que
nem todo o DNA humano foi seqüenciado. Estimativas atuais concluem que
apenas cerca de 2% do material genético humano é composto de genes,
enquanto que a maior parte parece não conter instruções para a formação
de proteínas, e existe provavelmente por razões estruturais. Muito pouco
dessa maior parte do material genético tem sua seqüência conhecida.
Por limitações tecnológicas, partes do DNA que
possuem muitas repetições de bases nitrogenadas também ainda não foram
totalmente seqüenciadas. Essas partes incluem, por exemplo, os
centrômeros e os telômeros dos cromossomos.
De todos os genes que tiveram sua seqüência determinada, aproximadamente 50% codificam para proteínas de função conhecida.
Apesar dessas lacunas, a conclusão do genoma já
está facilitando o desenvolvimento de fármacos muito mais potentes,
assim como a compreensão de diversas doenças genéticas humanas.
Do mesmo modo que uma fábrica pode ser
multiplicada pela construção de várias filiais, também as células se
dividem e produzem cópias de si mesmas.
Há dois tipos de divisão celular: mitose e meiose.
Na mitose, a
divisão de uma “célula-mãe” duas “células-filhas” geneticamente
idênticas e com o mesmo número cromossômico que existia na célula-mãe. Uma célula n produz duas células n, uma célula 2n produz duas células 2n etc. Trata-se de uma divisão equacional.
Já na meiose, a divisão de uma “célula-mãe” 2n gera “células-filhas” n, geneticamente diferentes. Neste caso, como uma célula 2n produz quatro células n, a divisão é chamada reducional.
A interfase – A fase que precede a mitose
É impossível imaginar a multiplicação de uma
fabrica, de modo que todas as filiais fossem extremamente semelhantes a
matriz, com cópias fieis de todos os componentes, inclusive dos
diretores? Essa, porém, no caso da maioria das células, é um
acontecimento rotineiro. A mitose corresponde a criação de uma cópia da
fabrica e sua meta é a duplicação de todos os componentes.
A principal atividade da célula, antes de se
dividir, refere-se a duplicação de seus arquivos de comando, ou seja, à
reprodução de uma cópia fiel dos dirigentes que se encontram no núcleo.
A interfase é o período que precede qualquer divisão celular, sendo de intensa atividade metabólica.
Nesse período, há a preparação para a divisão celular, que envolve a
duplicação da cromatina, material responsável pelo controle da atividade
da célula. Todas as informações existentes ao longo da molécula de DNA
são passadas para a cópia, como se correspondessem a uma cópia
fotográfica da molécula original. Em pouco tempo, cada célula formada da
divisão receberá uma cópia exata de cada cromossomo da célula se
dividiu.
As duas cópias de cada cromossomo permanecem
juntas por certo tempo, unidas pelo centrômero comum, constituindo duas
cromátides de um mesmo cromossomo. Na interfase, os centríolos também se
duplicam.
A interfase e a Duplicação do DNA
Houve época em que se falava que a interfase era o
período de “repouso” da célula. Hoje, sabemos, que na realidade a
interfase é um período de intensa atividade metabólica no ciclo celular:
é nela que se dá a duplicação do DNA, crescimento e síntese. Costuma-se dividir a interfase em três períodos distintos: G1, S e G2.
O intervalo de tempo em que ocorre a duplicação do DNA foi denominado de S (síntese) e o período que antecede é conhecido como G1 (G1 provém do inglês gap, que significa “intervalo”). O período que sucede o S é conhecido como G2.

O ciclo celular todo, incluindo a interfase (G1,
S, G2) e a mitose (M) – prófase, metáfase, anáfase e telófase – pode ser
representado em um gráfico no qual se coloca a a quantidade da DNA na
ordenada (y) e o tempo na abscissa (x). Vamos supor que a célula que vai
se dividir tenha, no período G1, uma quantidade 2C de DNA (C é uma
unidade arbitrária). O gráfico da variação de DNA, então, seria
semelhante ao da figura abaixo.

Nas células, existe uma espécie de
“manual de verificação de erros” que é utilizado em algumas etapas do
ciclo celular e que é relacionado aos pontos de checagem. Em cada ponto
de checagem a célula avalia se é possível avançar ou se é necessário
fazer algum ajuste, antes de atingir a fase seguinte. Muitas vezes, a
escolha é simplesmente cancelar o processo ou até mesmo conduzir a
célula à morte.
A mitose é um processo contínuo de divisão
celular, mas, por motivos didáticos, para melhor compreendê-la, vamos
dividi-la em fases: prófase, metáfase, anáfase e telófase. Alguns autores costumam citar uma quinta fase – a prometáfase – intermediária entre a prófase e a metáfase. O final da mitose, com a separação do citoplasma, é chamado de citocinese.
Prófase – Fase de início (pro = antes)
- Os cromossomos começam a ficar visíveis devido à espiralação.
- O nucléolo começa a desaparecer.
- Organiza-se em torno do núcleo um conjunto de fibras (nada mais são do que microtúbulos) originadas a partir dos centrossomos, constituindo o chamado fuso de divisão (ou fuso mitótico).
Embora os centríolos participem da divisão, não é
deles que se originam as fibras do fuso. Na mitose em célula animal, as
fibras que se situam ao redor de cada par de centríolos opostas ao fuso
constituem o áster (do grego, aster = estrela).
- O núcleo absorve água, aumenta de volume e a carioteca se desorganiza.
- No final da prófase, curtas fibras do fuso, provenientes do centrossomos, unem-se aos centrômeros. Cada uma das cromátides-irmãs fica ligada a um dos pólos da célula.
Note que os centrossomos ainda estão
alinhados na região equatorial da célula, o que faz alguns autores
designarem essa fase de prometáfase.

A formação de um novo par de centríolos é
iniciada na fase G1, continua na fase S e na fase G2 a duplicação é
completada. No entanto, os dois pares de centríolos permanecem reunidos
no mesmo centrossomo. Ao iniciar a prófase, o centrossomo parte-se em
dois e cada par de centríolos começa a dirigir-se para pólos opostos da
célula que irá entrar em divisão.

- Os cromossomos atingem o máximo em espiralação, encurtam e se localizam na região equatorial da célula.
- No finalzinho da metáfase e início da anáfase ocorre a duplicação dos centrômeros.


Anáfase – Fase do deslocamento (ana indica movimento ao contrário)
- As fibras do fuso começam a encurtar. Em conseqüência, cada lote de cromossomos-irmãos é puxado para os pólos opostos da célula.
Como cada cromátide passa a ser um novo cromossomo, pode-se considerar que a célula fica temporariamente tetraplóide.


- Os cromossomos iniciam o processo de desespirilação.
- Os nucléolos reaparecem nos novos núcleos celulares.
- A carioteca se reorganiza em cada núcleo-filho.
- Cada dupla de centríolos já se encontra no local definitivo nas futuras células-filhas.

Citocinese – Separando as células
A partição em duas copias é chamada de citocinese
e ocorre, na célula animal, de fora para dentro, isto é, como se a
célula fosse estrangulada e partida em duas (citocinese centrípeta). Há
uma distribuição de organelas pelas duas células-irmãs. Perceba que a
citocinese é, na verdade a divisão do citoplasma. Essa divisão pode ter
início já na anáfase, dependendo da célula.


Na mitose de células de vegetais superiores,
basicamente duas diferenças podem ser destacadas, em comparação com que
ocorre na mitose da célula animal:
-
A mitose ocorre sem centríolos. A partir de certos locais, correspondentes ao centrossomos, irradiam-se as fibras do fuso. Uma vez que não há centríolos, então não existe áster. Por esse motivo, diz-se que a mitose em células vegetais é anastral (do grego, an = negativo);
-
A citocinese é centrífuga, ocorre do centro para a periferia da célula. No início da telófase forma-se o fragmoplasmo, um conjunto de microtúbulos protéicos semelhantes aos do fuso de divisão. Os microtúbulos do fragmoplasto funcionam como andaimes que orientam a deposição de uma placa celular mediana semelhante a um disco, originada de vesículas fundidas do sistema golgiense. Progressivamente, a placa celular cresce em direção à periferia e, ao mesmo tempo, no interior da vesícula, ocorre a deposição de algumas substâncias, entre elas, pectina e hemicelulose, ambos polissacarídeos. De cada lado da placa celular, as membranas fundidas contribuem para a formação, nessa região, das membranas plasmáticas das duas novas células e que acabam se conectando com a membrana plasmática da célula-mãe. Em continuação à formação dessa lamela média, cada célula-filha, deposita uma parede celulósica primária, do lado de fora da membrana plasmática. A parede primária acaba se estendendo por todo o perímetro da célula. Simultaneamente a parede celulósica primária da célula-mãe é progressivamente desfeita, o que permite o crescimento de cada célula-filha, cada qual dotada, agora, de uma nova parede primária. Então, se pudéssemos olhar essa região mediana de uma das células, do citoplasma para fora, veríamos, inicialmente, a membrana plasmática, em seguida a parede celulósica primária e, depois, a lamela média. Eventualmente, uma parede secundária poderá ser depositada entre a membrana plasmática e a parede primária.

A mitose serve para...
A mitose é um tipo de divisão muito freqüente entre os organismos da Terra atual. Nos unicelulares, serve à reprodução assexuada e à multiplicação dos organismos. Nos pluricelulares, ela repara tecidos lesados, repões células que normalmente morrem e também está envolvida no crescimento.
No homem, a pele, a medula óssea e o revestimento intestinal são locais onde a mitose é freqüente. Nem todas as células do homem, porém, são capazes de realizar mitose.
Neurônios e célula musculares são dois tipos celulares altamente
especializados em que não ocorre esse tipo de divisão (ocorre apenas na
fase embrionária). Nos vegetais, a mitose ocorre em locais onde existem
tecidos responsáveis pelo crescimento, por exemplo, na ponta de raízes,
na ponta de caules e nas gemas laterais. Serve também para produzir
gametas, ao contrário do que ocorre nos animais, em que a meiose é o
processo de divisão mais diretamente associado à produção das células
gaméticas.
Como sabemos, a interfase é um período de intensa
atividade metabólica e de maior duração do ciclo celular. Células
nervosas e musculares, que não se dividem por mitose, mantêm-se
permanentemente na interfase, estacionadas no período chamado G0.
Nas células que se dividem ativamente, a
interfase é seguida da mitose, culminando na citocinese. Sabe-se que a
passagem de uma fase para outra é controlada por fatores de regulação - de modo geral protéicos – que atuam nos chamados pontos de checagem do ciclo celular. Dentre essas proteínas, se destacam as ciclinas, que controlam a passagem da fase G1 para a fase S e da G2 para a mitose.
Se em algumas dessas fases houver alguma
anomalia, por exemplo, algum dano no DNA, o ciclo é interrompido até que
o defeito seja reparado e o ciclo celular possa continuar. Caso
contrário, a célula é conduzida à apoptose (morte celular programada).
Outro ponto de checagem é o da mitose, promovendo
a distribuição correta dos cromossomos pelas células-filhas. Perceba
que o ciclo celular é perfeitamente regulado, está sob controle de
diversos genes e o resultado é a produção e diferenciação das células
componentes dos diferentes tecidos do organismo. Os pontos de checagem
correspondem, assim, a mecanismos que impedem a formação de células
anômalas.

A origem das células cancerosas está associada a anomalias na regulação do ciclo celular e à perda de controle da mitose.
Alterações do funcionamento de genes controladores do ciclo celular, em
decorrência de mutações, são relacionados ao surgimento de um câncer.
Duas classes de genes, os proto-onco-genes e os genes supressores de
tumor são os mais diretamente relacionados à regulação do ciclo celular.
Os proto-oncogenes são responsáveis pela produção de proteínas que
atuam na estimulação do ciclo celular, enquanto os genes supressores de
tumor são responsáveis pela produção de proteínas que atuam inibindo o
ciclo celular.
Dizendo de outro modo:
Os proto-oncogenes,
quando ativos, estimulam a ocorrência de divisão celular e os genes
supressores de tumor, quando ativos, inibem a ocorrência de divisão
celular. O equilíbrio na atuação desses dois grupos de genes resulta no
perfeito funcionamento do ciclo celular.
Mutações nos proto-oncogenes os transformam em
oncogenes ( genes causadores de câncer). As que afetam os genes
supressores de tumor perturbam o sistema inibidor e o ciclo celular fica
desregulado, promovendo a ocorrência desordenada de divisões celulares e
o surgimento de células cancerosas, que possuem as seguintes
características:
- são indiferenciadas, não contribuindo para a formação natural dos tecidos,
- seus núcleos são volumosos e com um número anormal de cromossomos;
- empilham-se sobre a outras em várias camadas, originando um aglomerado de células que forma um tumor. Se ficar restrito ao local de origem e for encapsulado, diz-se que o tumor é benigno, podendo ser removido;
- nos tumores malignos, ocorre a metástase, ou seja, as células cancerosas abandonam o local de origem, espalham-se por via sangüínea ou linfática, e invadem outros órgãos. Esse processo é acompanhado por uma angiogênese, que é a formação de inúmeros vasos sanguíneos responsáveis pela nutrição das células cancerosas.
Outra ocorrência envolvendo alterações do ciclo celular é relativa aos telômeros,
que são segmentos de moléculas de DNA com repetições de bases que atuam
como “capas protetoras” da extremidade dos cromossomos.
Em células humanas normais, a cada ciclo celular os telômeros são progressivamente encurtados, as extremidades dos cromossomos ficam cada vez mais curtas, até atingir um limite mínimo de tamanho incompatível com a vida da célula, paralisando-se as divisões celulares e sinalizando o fim da vida da célula. Em células cancerosas esse limite é transposto graças a atividade de uma enzima , a telomerase, que atua na reposição constante dos telômeros, mantendo-os sempre com o tamanho original, permitindo assim, que as células se dividam continuamente e se tornem praticamente “imortais”. |
 |
Diferentemente da mitose, em que uma célula
diplóide, por exemplo, se divide formando duas células também diplóides
(divisão equacional), a meiose é um tipo de divisão celular em que uma célula diplóide produz quatro células haplóides, sendo por este motivo uma divisão reducional.
Um fato que reforça o caráter reducional da
meiose é que, embora compreenda duas etapas sucessivas de divisão
celular, os cromossomos só se duplicam uma vez, durante a interfase –
período que antecede tanto a mitose como a meiose. No início da
interfase, os filamentos de cromatina não estão duplicados.
Posteriormente, ainda nesta fase, ocorre a duplicação, ficando cada
cromossomo com duas cromátides.
As várias fases da meiose
A redução do número cromossômico da célula é
importante fator para a conservação do lote cromossômico das espécies,
pois como a meiose formam-se gametas com a metade do lote cromossômico.
Quando da fecundação, ou seja, do encontro de dois gametas, o número de
cromossomos da espécie se restabelece.
Podemos estudar a meiose em duas etapas, separadas por um curto intervalo, chamado intercinese. Em cada etapa, encontramos as fases estudadas na mitose, ou seja, prófase, metáfase, anáfase e telófase.
Vamos supor uma célula 2n = 2 e estudar os eventos principais da meiose nessa célula.
Meiose I (Primeira Divisão Meiótica)
Prófase I –
É a etapa mais marcante da meiose. Nela ocorre o pareamento dos
cromossomos homólogos e pode acontecer um fenômeno conhecido como
crossing-over (também chamado de permuta)
Como a prófase I é longa, há uma seqüência de eventos que, para efeito de estudo, pode ser dividida nas seguintes etapas:
- Inicia-se a espiralação cromossômica. É a fase de leptóteno (leptós = fino), em que os filamentos cromossômicos são finos, pouco visíveis e já constituídos cada um por duas cromátides.

- Começa a atração e o pareamento dos cromossomos homólogos; é um pareamento ponto por ponto conhecido como sinapse (o prefixo sin provém do grego e significa união). Essa é a fase de zigóteno (zygós = par).

- A espiralação progrediu: agora, são bem visíveis as duas cromátides de cada homólogo pareado; como existem, então, quatro cromátides, o conjunto forma uma tétrade ou par bivalente. Essa é a fase de paquíteno (pakhús = espesso).

- Ocorrem quebras casuais nas cromátides e uma troca de pedaços entre as cromátides homólogas, fenômeno conhecido como crossing-over (ou permuta). Em seguida, os homólogos se afastam e evidenciam-se entre eles algumas regiões que estão ainda em contato. Essas regiões são conhecidas como quiasmas (qui corresponde à letra “x” em grego). Os quiasmas representam as regiões em que houve as trocas de pedaços. Essa fase da prófase I é o diplóteno (diplós = duplo).

- Os pares de cromátides fastam-se um pouco mais e os quiasmas parecem “escorregar” para as extremidades; a espiralação dos cromossomos aumenta. è a última fase da prófase I, conhecida por diacinese (dia = através; kinesis = movimento).

Enquanto acontecem esses eventos, os
centríolos, que vieram duplicado da interfase, migram para os pólos
opostos e organizam o fuso de divisão; os nucléolos desaparecem; a
carioteca se desfaz após o término da prófase I, prenunciando a
ocorrência da metáfase I.
Metáfase I – os cromossomos homólogos pareados se dispõem na região mediana da célula; cada cromossomo está preso a fibras de um só pólo.
Anáfase I
– o encurtamento das fibras do fuso separa os cromossomos homólogos,
que são conduzidos para pólos opostos da célula, não há separação das
cromátides-irmãs. Quando os cromossomos atingem os pólos, ocorre sua
desespiralação, embora não obrigatória, mesmo porque a segunda etapa da
meiose vem a seguir. Às vezes, nem mesmo a carioteca se reconstitui.
Telófase I
– no final desta fase, ocorre a citocinese, separando as duas
células-filhas haplóides. Segue-se um curto intervalo a intercinese, que
procede a prófase II.

Prófase II
– cada uma das duas células-filhas tem apenas um lote de cromossomos
duplicados. Nesta fase os centríolos duplicam novamente e as células em
que houve formação da carioteca, esta começa a se desintegrar.
Metáfase II - como na mitose, os cromossomos prendem-se pelo centrômero às fibras do fuso, que partem de ambos os pólos.
Anáfase II – Ocorre duplicação dos centrômeros, só agora as cromátides-irmãs separam-se (lembrando a mitose).
Telófase II e citocinese
– com o término da telófase II reorganizam-se os núcleos. A citocinese
separa as quatro células-filhas haplóides, isto é, sem cromossomos
homólogos e com a metade do número de cromossomos em relação à célula
que iniciou a meiose.

A principal conseqüência da meiose, sem dúvida, é o surgimento da diversidade entre os indivíduos que são produzidos na reprodução sexuada da espécie.
A relação existente entre meiose e variabilidade é baseada principalmente na ocorrência de crossing-over.
 |
O crossing é
um fenômeno que envolve cromátides homólogas. Consiste na quebra dessas
cromátides em certos pontos, seguida de uma troca de pedaços
correspondentes entre elas.
As trocas provocam o surgimento de
novas seqüências de genes ao longo dos cromossomos. Assim, se em um
cromossomo existem vários genes combinados segundo uma certa seqüência,
após a ocorrência do crossing a combinação pode não ser mais a mesma.
Então, quando se pensa no crossing, é comum analisar o que aconteceria,
por exemplo, quanto à combinação entre os genes alelos A e a e B e b no par de homólogos ilustrados na figura.
Nessa combinação o gene A e B encontram-se em um mesmo cromossomo, enquanto a e b estão no cromossomo homólogo. Se a distância de A e B for considerável, é grande a chance de ocorrer uma permuta. E, se tal acontecer, uma nova combinação gênica poderá surgir.
As combinações Ab e aB
são novas. São recombinações gênicas que contribuem para a geração de
maior variabilidade nas células resultantes da meiose. Se pensarmos na
existência de três genes ligados em um mesmo cromossomo (A, b e C, por
exemplo), as possibilidades de ocorrência de crossings dependerão da
distância em que os genes se encontram – caso estejam distantes, a
variabilidade produzida será bem maior.
|
Outro processo que conduz ao surgimento de
variabilidade na meiose é a segregação independente dos cromossomos.
Imaginando-se que uma célula com dois pares de cromossomos homólogos (A e
a, B e b), se divida por meiose, as quatro células resultantes ao final
da divisão poderão ter a seguinte constituição cromossômica: (a e b), (a e B), (A e b) e (A e B).
A variabilidade genética existente entre os
organismos das diferentes espécies é muito importante para a ocorrência
da evolução biológica. Sobre essa variabilidade é que atua a seleção
natural, favorecendo a sobrevivência de indivíduos dotados de
características genéticas adaptadas ao meio. Quanto maior a
variabilidade gerada na meiose, por meio de recombinação gênica
permitida pelo crossing-over, maiores as chances para a ação seletiva do
meio.

Na meiose a variação da quantidade
de DNA pode ser representada como no gráfico ao lado, partindo-se, por
exemplo, de uma célula que tenha uma quantidade 2C de DNA em G1
Gametogênese
Gametogênese é o processo pelo qual os gametas são produzidos nos organismos dotados de reprodução sexuada. Nos animais, a gametogênese acontece nas gônadas, órgãos que também produzem os hormônios sexuais, que determinam as características que diferenciam os machos das fêmeas.
O evento fundamental da gametogênese é a meiose, que reduz à metade a quantidade de cromossomos das células, originando células haplóides. Na fecundação, a fusão de dois gametas haplóides reconstitui o número diplóide característico de cada espécie.
Em alguns raros casos, não acontece meiose durante a formação dos gametas. Um exemplo bastante conhecido é o das abelhas: se um óvulo não for fecundado por nenhum espermatozóide, irá se desenvolver por mitoses consecutivas, originando um embrião em que todas as células são haplóides. Esse embrião haplóide formará um indivíduo do sexo masculino. O desenvolvimento de um gameta sem que haja fecundação chama-se partenogênese. Se o óvulo for fecundado, o embrião 2n irá originar uma fêmea.
Em linhas gerais, a gametogênese masculina (ou espermatogênese) e a gametogênese feminina (ovogênese ou ovulogênese) seguem as mesmas etapas.
A Espermatogênese
Processo que ocorre nos testículos, as gônadas masculinas.
Secretam a testosterona, hormônio sexual responsável pelo aparecimento
das características sexuais masculinas: aparecimento da barba e dos
pêlos corporais em maior quantidade, massa muscular mais desenvolvida,
timbre grave da voz, etc.
As células dos testículos estão organizadas ao redor dos túbulos seminíferos, nos quais os espermatozóides são produzidos. A testosterona é secretada pelas células intersticiais. Ao redor dos túbulos seminíferos, estão as células de Sertoli, responsáveis pela nutrição e pela sustentação das células da linhagem germinativa, ou seja, as que irão gerar os espermatozóides.
As células dos testículos estão organizadas ao redor dos túbulos seminíferos, nos quais os espermatozóides são produzidos. A testosterona é secretada pelas células intersticiais. Ao redor dos túbulos seminíferos, estão as células de Sertoli, responsáveis pela nutrição e pela sustentação das células da linhagem germinativa, ou seja, as que irão gerar os espermatozóides.

Nos mamíferos, geralmente os testículos ficam fora da cavidade abdominal, em uma bolsa de pele chamada bolsa escrotal. Dessa forma, a temperatura dos testículos permanece aproximadamente 1° C inferior à temperatura corporal, o que é ideal para a espermatogênese.
A espermatogênese divide-se em quatro fases:
Fase de proliferação ou de multiplicação:
Tem início durante a vida intra-uterina, antes mesmo do nascimento do
menino, e se prolonga praticamente por toda a vida. As células
primordiais dos testículos, diplóides, aumentam em quantidade por
mitoses consecutivas e formam as espermatogônias .
|
 |
Fase de crescimento: Um pequeno aumento no volume do citoplasma das espermatogônias as converte em espermatócitos de primeira ordem, também chamados espermatócitos primários ou espermatócitos I, também diplóides. |
|
Fase de maturação: Também é rápida, nos machos, e corresponde ao período de ocorrência da meiose. Depois da primeira divisão meiótica, cada espermatócito de primeira ordem origina dois espermatócitos de segunda ordem (espermatócitos secundários ou espermatócitos II). Como resultam da primeira divisão da meiose, já são haplóides, embora possuam cromossomos duplicados. Com a ocorrência da segunda divisão meiótica, os dois espermatócitos de segunda ordem originam quatro espermátides haplóides. |
|
Espermiogênese:
É o processo que converte as espermátides em espermatozóides, perdendo
quase todo o citoplasma. As vesículas do complexo de Golgi fundem-se,
formando o acrossomo, localizado na extremidade anterior dos
espermatozóides. O acrossomo contém enzimas que perfuram as membranas do
óvulo, na fecundação.
Os centríolos migram para a região
imediatamente posterior ao núcleo da espermátide e participam da
formação do flagelo, estrutura responsável pela movimentação dos
espermatozóides. grande quantidade de mitocôndrias, responsáveis pela
respiração celular e pela produção de ATP, concentram-se na região entre
a cabeça e o flagelo, conhecida como peça intermediária.
|
Gametogênese
Gametogênese é o processo pelo qual os gametas são produzidos nos organismos dotados de reprodução sexuada. Nos animais, a gametogênese acontece nas gônadas, órgãos que também produzem os hormônios sexuais, que determinam as características que diferenciam os machos das fêmeas.
O evento fundamental da gametogênese é a meiose, que reduz à metade a quantidade de cromossomos das células, originando células haplóides. Na fecundação, a fusão de dois gametas haplóides reconstitui o número diplóide característico de cada espécie.
Em alguns raros casos, não acontece meiose durante a formação dos gametas. Um exemplo bastante conhecido é o das abelhas: se um óvulo não for fecundado por nenhum espermatozóide, irá se desenvolver por mitoses consecutivas, originando um embrião em que todas as células são haplóides. Esse embrião haplóide formará um indivíduo do sexo masculino. O desenvolvimento de um gameta sem que haja fecundação chama-se partenogênese. Se o óvulo for fecundado, o embrião 2n irá originar uma fêmea.
Em linhas gerais, a gametogênese masculina (ou espermatogênese) e a gametogênese feminina (ovogênese ou ovulogênese) seguem as mesmas etapas.
A Espermatogênese
Processo que ocorre nos testículos, as gônadas masculinas.
Secretam a testosterona, hormônio sexual responsável pelo aparecimento
das características sexuais masculinas: aparecimento da barba e dos
pêlos corporais em maior quantidade, massa muscular mais desenvolvida,
timbre grave da voz, etc.
As células dos testículos estão organizadas ao redor dos túbulos seminíferos, nos quais os espermatozóides são produzidos. A testosterona é secretada pelas células intersticiais. Ao redor dos túbulos seminíferos, estão as células de Sertoli, responsáveis pela nutrição e pela sustentação das células da linhagem germinativa, ou seja, as que irão gerar os espermatozóides.
As células dos testículos estão organizadas ao redor dos túbulos seminíferos, nos quais os espermatozóides são produzidos. A testosterona é secretada pelas células intersticiais. Ao redor dos túbulos seminíferos, estão as células de Sertoli, responsáveis pela nutrição e pela sustentação das células da linhagem germinativa, ou seja, as que irão gerar os espermatozóides.
Nos mamíferos, geralmente os testículos ficam fora da cavidade abdominal, em uma bolsa de pele chamada bolsa escrotal. Dessa forma, a temperatura dos testículos permanece aproximadamente 1° C inferior à temperatura corporal, o que é ideal para a espermatogênese.
A espermatogênese divide-se em quatro fases:
Fase de proliferação ou de multiplicação:
Tem início durante a vida intra-uterina, antes mesmo do nascimento do
menino, e se prolonga praticamente por toda a vida. As células
primordiais dos testículos, diplóides, aumentam em quantidade por
mitoses consecutivas e formam as espermatogônias .
|
|
Fase de crescimento: Um pequeno aumento no volume do citoplasma das espermatogônias as converte em espermatócitos de primeira ordem, também chamados espermatócitos primários ou espermatócitos I, também diplóides. |
|
Fase de maturação: Também é rápida, nos machos, e corresponde ao período de ocorrência da meiose. Depois da primeira divisão meiótica, cada espermatócito de primeira ordem origina dois espermatócitos de segunda ordem (espermatócitos secundários ou espermatócitos II). Como resultam da primeira divisão da meiose, já são haplóides, embora possuam cromossomos duplicados. Com a ocorrência da segunda divisão meiótica, os dois espermatócitos de segunda ordem originam quatro espermátides haplóides. |
|
Espermiogênese:
É o processo que converte as espermátides em espermatozóides, perdendo
quase todo o citoplasma. As vesículas do complexo de Golgi fundem-se,
formando o acrossomo, localizado na extremidade anterior dos
espermatozóides. O acrossomo contém enzimas que perfuram as membranas do
óvulo, na fecundação.
Os centríolos migram para a região
imediatamente posterior ao núcleo da espermátide e participam da
formação do flagelo, estrutura responsável pela movimentação dos
espermatozóides. grande quantidade de mitocôndrias, responsáveis pela
respiração celular e pela produção de ATP, concentram-se na região entre
a cabeça e o flagelo, conhecida como peça intermediária.
|
Nos ovários, encontram-se agrupamentos celulares chamados folículos ovarianos de Graff,
onde estão as células germinativas, que originam os gametas, e as
células foliculares, responsáveis pela manutenção das células
germinativas e pela produção dos hormônios sexuais femininos.
Nas mulheres, apenas um folículo ovariano entra em maturação a cada ciclo menstrual, período compreendido entre duas menstruações consecutivas e que dura, em média, 28 dias. Isso significa que, a cada ciclo, apenas um gameta torna-se maduro e é liberado no sistema reprodutor da mulher.
Os ovários alternam-se na maturação dos seus folículos, ou seja, a cada ciclo menstrual, a liberação de um óvulo, ou ovulação, acontece em um dos dois ovários.
Nas mulheres, apenas um folículo ovariano entra em maturação a cada ciclo menstrual, período compreendido entre duas menstruações consecutivas e que dura, em média, 28 dias. Isso significa que, a cada ciclo, apenas um gameta torna-se maduro e é liberado no sistema reprodutor da mulher.
Os ovários alternam-se na maturação dos seus folículos, ou seja, a cada ciclo menstrual, a liberação de um óvulo, ou ovulação, acontece em um dos dois ovários.


A ovogênese é dividida em três etapas:
 |
Fase de multiplicação ou de proliferação:
É uma fase de mitoses consecutivas, quando as células germinativas
aumentam em quantidade e originam ovogônias. Nos fetos femininos
humanos, a fase proliferativa termina por volta do final do primeiro
trimestre da gestação. Portanto, quando uma menina nasce, já possui em
seus ovários cerca de 400 000 folículos de Graff. É uma quantidade
limitada, ao contrário dos homens, que produzem espermatogônias durante
quase toda a vida.
|
Fase de crescimento: Logo que são formadas, as ovogônias iniciam a primeira divisão da meiose, interrompida na prófase I. Passam, então, por um notável crescimento, com aumento do citoplasma e grande acumulação de substâncias nutritivas. Esse depósito citoplasmático de nutrientes chama-se vitelo, e é responsável pela nutrição do embrião durante seu desenvolvimento. Terminada a fase de crescimento, as ovogônias transformam-se em ovócitos primários (ovócitos de primeira ordem ou ovócitos I). Nas mulheres, essa fase perdura até a puberdade, quando a menina inicia a sua maturidade sexual. |
|
Fase de maturação: Dos 400 000 ovócitos primários, apenas 350 ou 400 completarão sua transformação em gametas maduros, um a cada ciclo menstrual. A fase de maturação inicia-se quando a menina alcança a maturidade sexual, por volta de 11 a 15 anos de idade. Quando o ovócito primário completa a primeira divisão da meiose, interrompida na prófase I, origina duas células. Uma delas não recebe citoplasma e desintegra-se a seguir, na maioria das vezes sem iniciar a segunda divisão da meiose. É o primeiro corpúsculo (ou glóbulo) polar. A outra célula, grande e rica em vitelo, é o ovócito secundário (ovócito de segunda ordem ou ovócito II). Ao sofrer, a segunda divisão da meiose, origina o segundo corpúsculo polar, que também morre em pouco tempo, e o óvulo, gameta feminino, célula volumosa e cheia de vitelo. |
Na gametogênese feminina, a divisão meiótica é desigual porque não reparte igualmente o citoplasma entre as células-filhas. Isso permite que o óvulo formado seja bastante rico em substâncias nutritivas.
Na maioria das fêmeas de mamíferos, a segunda divisão da meiose só acontece caso o gameta seja fecundado. Curiosamente, o verdadeiro gameta dessas fêmeas é o ovócito II, pois é ele que se funde com o espermatozóide.
Para que surja um novo indivíduo, os gametas fundem-se aos pares, um masculino e outro feminino, que possuem papéis diferentes na formação do descendente. Essa fusão é a fecundação ou fertilização.
Ambos trazem a mesma quantidade haplóide de cromossomos, mas apenas os gametas femininos possuem nutrientes, que alimentam o embrião durante o seu desenvolvimento. Por sua vez, apenas os gametas masculinos são móveis, responsáveis pelo encontro que pode acontecer no meio externo (fecundação externa) ou dentro do corpo da fêmea (fecundação interna). Excetuando-se muitos dos artrópodes, os répteis, as aves e os mamíferos, todos os outros animais possuem fecundação externa, que só acontece em meio aquático.

Quando a fecundação é externa, tanto os machos quanto as fêmeas produzem gametas em grande quantidade, para compensar a perda que esse ambiente ocasiona. Muitos gametas são levados pelas águas ou servem de alimentos para outros animais. Nos animais dotados de fecundação interna, as fêmeas produzem apenas um ou alguns gametas por vez, e eles encontram-se protegidos dentro do sistema reprodutor.
 |
Além da membrana plasmática, o óvulo possui outro revestimento mais externo, a membrana vitelínica.
Quando um espermatozóide faz contato com a membrana vitelínica, a
membrana do acrossomo funde-se à membrana do espermatozóide (reação
acrossômica), liberando as enzimas presentes no acrossomo.
As enzimas do acrossomo dissolvem a membrana vitelínica e abrem caminho para a penetração do espermatozóide. Com a fusão da membrana do espermatozóide com a membrana do óvulo, o núcleo do espermatozóide penetra no óvulo. Nesse instante, a membrana do óvulo sofre alterações químicas e elétricas, transformando-se na membrana de fertilização, que impede a penetração de outros espermatozóides. No interior do óvulo, o núcleo do espermatozóide, agora chamado pró-núcleo masculino, funde-se com o núcleo do óvulo, o pró-núcleo feminino. Cada pró-núcleo traz um lote haplóide de cromossomos, e a fusão resulta em um lote diplóide, o zigoto. Nessa célula, metade dos cromossomos tem origem paterna e metade, origem materna. |
Os ácidos nucléicos são macromoléculas de
natureza química, formadas por nucleotídeos, grupamento fosfórico
(fosfato), glicídio (monossacarídeo / pentoses) e uma base nitrogenada,
compondo o material genético contido nas células de todos os seres
vivos.

Presentes no núcleo dos eucariotos e dispersos no hialoplasma dos procariotos, os ácidos nucléicos podem ser de dois tipos: ácido desoxirribonucléico (DNA) e ácido ribonucléico (RNA),
ambos relacionados ao mecanismo de controle metabólico celular
(funcionamento da célula) e transmissão hereditária das características.
Os Ácidos Nucléicos: DNA e RNA
O DNA se diferencia do RNA por possuir o açúcar desoxirribose e os nucleotídeos adenina, citosina, guanina e timina. No RNA, o açúcar é a ribose e os nucleotídeos são adenina, citosina, guanina e uracila (a uracila entra no lugar da timina).

A partir de experimentos feitos por vários
pesquisadores e utilizando os resultados da complexa técnica de difração
com raios X, Watson e Crick concluíram que, no DNA, as cadeias
complementares são helicoidais, sugerindo a idéia de uma escada
retorcida.
Nessa escada, os corrimãos são formados por
fosfatos e desoxirribose, enquanto os degraus são constituídos pelos
pares de bases nitrogenadas.


Os átomos de carbono das moléculas de ribose e desoxirribose são numerados conforme a figura abaixo. Observe que os carbonos do açúcar são numerados com uma linha (‘) a fim de distingui-los dos outros carbonos do nucleotídeo.

Em cada fita do DNA, o “corrimão” é formado por
ligações entre moléculas de açúcar e radicais fosfato. Note que o
radical fosfato se liga ao carbono 3’ de um açúcar e ao carbono 5’ do
seguinte.

As duas cadeias de nucleotídeos do DNA são unidas uma à outra por ligações chamadas de pontes de hidrogênio, que se formam entre as bases nitrogenadas de cada fita.
O pareamento de bases ocorre de maneira precisa: uma base púrica se liga a uma pirimídica – adenina (A) de uma cadeia pareia com a timina (T) da outra e guanina (G) pareia com citosina (C).

O DNA controla toda a atividade celular. Ele possui a
“receita” para o funcionamento de uma célula. Toda vez que uma célula
se divide, a “receita” deve ser passada para as células-filhas. Todo o
“arquivo” contendo as informações sobre o funcionamento celular precisa
ser duplicado para que cada célula-filha receba o mesmo tipo de
informação que existe na célula-mãe. Para que isso ocorra, é fundamental
que o DNA sofra “auto-duplicação”.
O modelo estrutural do DNA proposto por Watson e
Crick explica a duplicação dos genes: as duas cadeias do DNA se separam e
cada uma delas orienta a fabricação de uma metade complementar.
O experimento dos pesquisadores Meselson e Stahl
confirmou que a duplicação do DNA é semiconservativa, isto é, que metade
da molécula original se conserva íntegra em cada uma das duas
moléculas-filhas.

No processo de duplicação do DNA, as pontes de
hidrogênio entre as bases se rompem e as duas cadeias começam a se
separar. À medida que as bases vão sendo expostas, nucleotídeos que
vagam pelo meio ao redor vão se unindo a elas, sempre respeitando a
especificidade de emparelhamento: A com T, T com A, C com G e G com C.
Uma vez ordenados sobre a cadeia que está que está servindo de modelo,
os nucleotídeos se ligam em seqüência e formam uma cadeia complementar
dobre cada uma das cadias da molécula original. Assim, uma molécula de
DNA reproduz duas moléculas idênticas a ela.

A ação da enzima DNA polimerase
Diversos aspectos da duplicação do DNA já foram
desvendados pelos cientistas. Hoje, sabe-se que há diversas enzimas
envolvidas nesse processo. Certas enzimas desemparelham as duas cadeias
de DNA, abrindo a molécula. Outras desenrolam a hélice dupla, e há,
ainda, aquelas que unem os nucleotídeos entre si. A enzima que promove a
ligação dos nucleotídeos é conhecida como DNA polimerase, pois sua
função é construir um polímero (do grego poli, muitas, e meros, parte)
de nucleotídeos.

DNA polimerase
O material genético representado pelo DNA contém
uma mensagem em código que precisa ser decifrada e traduzida em
proteínas, muitas das quais atuarão nas reações metabólicas da célula. A
mensagem contida no DNA deve, inicialmente, ser passada para moléculas
de RNA que, por sua vez, orientarão a síntese de proteínas. O controle
da atividade celular pelo DNA, portanto, é indireto e ocorre por meio da
fabricação de moléculas de RNA, em um processo conhecido como
transcrição.
RNA: Uma Cadeia (Fita) Simples
As moléculas de RNA são constituídas por uma seqüência de ribonucleotídeos, formando uma cadeia (fita) simples.

Existem três tipos básicos de RNA, que diferem um do outro no peso molecular: o RNA ribossômico, representado por RNAr (ou rRNA), o RNA mensageiro, representado por RNAm (ou mRNA) e o RNA transportador, representado por RNAt (ou tRNA).
- O RNA ribossômico é o de maior peso molecular e constituinte majoritário do ribossomo, organóide relacionado à síntese de proteínas na célula.
- O RNA mensageiro é o de peso molecular intermediário e atua conjuntamente com os ribossomos na síntese protéica.
- O RNA transportador é o mais leve dos três e encarregado de transportar os aminoácidos que serão utilizados na síntese de proteínas.

A síntese de RNA (mensageiro, por exemplo) se
inicia com a separação das duas fitas de DNA. Apenas uma das fitas do
DNA serve de molde para a produção da molécula de RNAm. A outra fita não
é transcrita. Essa é uma das diferenças entre a duplicação do DNA e a
produção do RNA.

As outras diferenças são:
- os nucleotídeos utilizados possuem o açúcar ribose no lugar da desoxirribose;
- há a participação de nucleotídeos de uracila no lugar de nucleotídeos de timina. Assim, se na fita de DNA que está sendo transcrita aparecer adenina, encaminha-se para ela um nucleotídeo complementar contendo uracila;
Imaginando um segmento hipotético de um filamento de DNA com a seqüência de bases:
DNA- ATGCCGAAATTTGCG
O segmento de RNAm formado na transcrição terá a seqüência de bases:
RNA- UACGGCUUUAAACGC
Em uma célula eucariótica, o RNAm produzido
destaca-se de seu molde e, após passar por um processamento, atravessa a
carioteca e se dirige para o citoplasma, onde se dará a síntese
protéica. Com o fim da transcrição, as duas fitas de DNA seu unem
novamente, refazendo-se a dupla hélice.

A mensagem genética contida no DNA é formada por um alfabeto de quatro letras que correspondem aos quatro nucleotídeos: A, T, C e G.
Com essas quatros letras é preciso formar “palavras” que possuem o
significado de “aminoácidos”. Cada proteína corresponde a uma “frase”
formada pelas “palavras”, que são os aminoácidos. De que maneira apenas
quatro letras do alfabeto do DNA poderiam ser combinadas para
corresponder a cada uma das vinte “palavras” representadas pelos vinte
aminoácidos diferentes que ocorrem nos seres vivos.

Uma proposta brilhante sugerida por vários pesquisadores, e depois confirmada por métodos experimentais, foi a de que cada três letras (uma trinca de bases) do DNA corresponderia uma “palavra”, isto é, um aminoácido. Nesse caso, haveria 64 combinações possíveis de três letras, o que seria mais do que suficiente para codificar os vinte tipos diferentes de aminoácidos (matematicamente,
utilizando o método das combinações seriam, então, 4 letras combinadas 3
a 3, ou seja, 43 = 64 combinações possíveis).
O código genético do DNA se expressa por trincas de bases, que foram denominadas códons. Cada códon, formado por três letras, corresponde a um certo aminoácido.
A correspondência entre o trio de bases do DNA, o
trio de bases do RNA e os aminoácidos por eles especificados constitui
uma mensagem em código que passou a ser conhecida como “código
genético”.
Mas, surge um problema. Como são vinte os
diferentes aminoácidos, há mais códons do que tipos de aminoácidos!
Deve-se concluir, então, que há aminoácidos que são especificados por
mais de um códon, o que foi confirmado. A tabela abaixo, especifica os códons de RNAm que podem ser formados e os correspondentes aminoácidos que especificam.

Dizemos que o código genético é universal, pois
em todos os organismos da Terra atual ele funciona da mesma maneira,
quer seja em bactérias, em uma cenoura ou no homem.
O códon AUG, que codifica para o aminoácido
metionina, também significa início de leitura, ou seja, é um códon que
indica aos ribossomos que é por esse trio de bases qe deve ser iniciada a
leitura do RNAm.
Note que três códons não especificam nenhum aminoácido. São os códons UAA, UAG e UGA, chamados de códons e parada durante a “leitura” (ou stop códons) do RNA pelos ribossomos, na síntese protéica.
Diz-se que o código genético é degenerado
porque cada “palavra” (entenda-se aminoácido) pode ser especificada por
mais de uma trinca.
Tradução é o nome utilizado para designar o
processo de síntese de proteínas. Ocorre no citoplasma com a
participação, entre outros, de RNA e de aminoácidos.
Quem participa da síntese de proteínas?
Cístron (gene) é o segmento de DNA que contém as informações para a síntese de um polipeptídeo ou proteína.
O RNA produzido que contém uma seqüência de bases nitrogenadas transcrita do DNA é um RNA mensageiro.

No citoplasma, ele será um dos componentes
participantes da síntese de proteínas, juntamente com outros dois tipos
de RNA, todos de fita simples e produzidos segundo o mesmo processo
descrito para o RNA mensageiro:
RNA ribossômico, RNAr.
Associando-se a proteínas, as fitas de RNAr formarão os ribossomos,
orgânulos responsáveis pela leitura da mensagem contida no RNA
mensageiro;
|

RNAs transportadores, RNAt.
Assim chamados porque serão os responsáveis pelo transporte de
aminoácidos até o local onde se dará a síntese de proteínas junto aos
ribossomos. São moléculas de RNA de fita simples, de pequeno tamanho,
contendo, cada uma, cerca de 75 a 85 nucleotídeos. Cada fita de RNAt
torce-se sobre si mesma, adquirindo o aspecto visto na figura abaixo.
Duas regiões se destacam em cada
transportador: uma é o local em que se ligará o aminoácido a ser
transportado e a outra corresponde ao trio de bases complementares
(chamado anticódon) do RNAt, que se encaixará no códon correspondente do
RNAm.
Anticódon é o trio de bases do RNAt, complementar do códon do RNAm.
|

Cada RNAt em solução liga-se a um determinado
aminoácido, formando-se uma molécula chamada aminoacil-RNAt, que
conterá, na extremidade correspondente ao anticódon, um trio de códon do
RNAm.
Para entendermos bem este processo, vamos admitir
que ocorra a síntese de um peptídeo contendo apenas sete aminoácidos, o
que se dará a partir da leitura de um RNAm contendo sete códons (21
bases hidrogenadas). A leitura (tradução) será efetuada por um ribossomo
que se deslocará ao longo do RNAm.


Esquematicamente na síntese proteica teríamos:
- Um RNAm, processado no núcleo, contendo sete códons (21 bases hidrogenadas) se dirige ao citoplasma.
- No citoplasma, um ribossomo se liga ao RNAm na extremidade correspondente ao início da leitura. Dois RNAt, carregando os seus respectivos aminoácidos (metionina e alanina), prendem-se ao ribossomo. Cada RNAt liga-se ao seu trio de bases (anticódon) ao trio de bases correspondentes ao códon do RNAm. Uma ligação peptídica une a metionina à alanina.
- O ribossomo se desloca ao longo do RNAm. O RNAt que carregava a metionina se desliga do ribossomo. O quarto RNAt, transportando o aminoácido leucina, une o seu anticódon ao códon correspondente do RNAm. Uma ligação peptídica é feita entre a leucina e a alanina.
- O ribossomo novamente se desloca. O RNAt que carregava a alanina se desliga do ribossomo. O quarto RNAt, transportando o aminoácido ácido glutâmico encaixa-se no ribossomo. Ocorre a união do anticódon desse RNAt com o códon correspondente do RNAm. Uma ligação peptídica une o ácido glutâmico à leucina.
- Novo deslocamento do ribossomo. O quinto RNAt, carregando a aminoácido glicina, se encaixa no ribossomo. Ocorre a ligação peptídica da glicina com o ácido glutâmico.
- Continua o deslocamento do ribossomo ao longo do RNAm. O sexto RNAt, carregando o aminoácido serina, se encaixa no ribossomo. Uma liogação peptídica une a serina à glicina.
- Fim do deslocamento do ribossomo. O último transportador , carregando o aminoácido triptofano, encaixa-se no ribossomo. Ocorre a ligação peptídica do triptofano com a serina. O RNAt que carrega o triptofano se separa do ribossomo. O mesmo ocorre com o transportador que portava a serina.
- O peptídeo contendo sete aminoácidos fica livre no citoplasma. Claro que outro ribossomo pode se ligar ao RNAm, reiniciando o processo de tradução, que resultará em um novo peptídio. Perceba, assim, que o RNAm contendo sete códons (21 bases nitrogenadas) conduziu a síntese de um peptídeo formado por sete aminoácidos.
Em algumas células, certas proteínas são
produzidas em grande quantidade. Por exemplo, a observação de glândulas
secretoras de certos hormônios de natureza protéica (que são liberados
para o sangue, indo atuar em outros órgãos do mesmo organismo) mostra,
em certos locais, uma fileira de ribossomos efetuando a leitura do mesmo
RNA mensageiro. Assim, grandes quantidades da mesma proteína são
produzidas.
Ao conjunto de ribossomos, atuando ao longo se um RNAm, dá-se o nome de polirribossomos.


Possíveis destinos das proteínas
O RNAm, ao sair do nucleo pode seguir dois destinos:
- Ser traduzido nos ribossomos do retículo endoplasmático rugoso - e esta proteína será exportada para fora da célula passando pelo Golgi e saindo por exocitose em forma de vesículas,
- Ou será traduzido nos ribossomas livres no citoplasma - esta proteína então permanecerá dentro da célula, executando alguma importante função. Ex. Dentro do Golgi, ou livre no citoplasma.

Síntese de proteínas no núcleo celular?
No núcleo de todas as células eucarióticas, a
produção de RNA se da a partir do molde de uma fita do DNA. Então, o RNA
sofre algumas alterações e segue para o citoplasma onde se associa aos
ribossomos para a formação das proteínas. Acreditava-se que a síntese de
proteínas (tradução) ocorresse somente no citoplasma, mas em recente
trabalho publicado na prestigiada revista Science foi demonstrado que os
elementos necessários à tradução se associam no núcleo, onde proteínas
seriam formadas. Além disso os pesquisadores constataram que as
estruturas responsáveis pela tradução estão em atividade no núcleo
celular.
Todos os dias as suas células produzem proteínas
que contêm aminoácidos em uma certa seqüência. Imagine, por exemplo, que
em um certo dia uma célula da epiderme de sua pele produza uma proteína
diferente. Suponha também que essa proteína seja uma enzima que atue em
uma reação química que leva a produção de um pigmento amarelo em vez do
pigmento normalmente encontrado na pele, a melanina. Essa célula se
multiplica e de repente aparece uma mancha amarelada em sua pele.
Provavelmente essa proteína poderá ter sofrido uma alteração em sua
seqüência de aminoácidos, tendo havido a substituição de um aminoácido
por outro, o que acarretou uma mudança em seu mecanismo de atuação e,
como conseqüência levou à produção de um pigmento de cor diferente.
Agora, como a seqüência de aminoácidos em uma proteína é determinada
pela ação de um certo gene que conduz à síntese do pigmento.

Essa alteração na seqüência de bases na molécula de DNA constituinte do gene é que se chama de mutação gênica.


O albinismo é causada por uma mutação na enzima
tirozinase que transforma o aminoácido tirozina em pigmento da pele, a
melanina. Esta doença ocorre em animais e nas plantas e é hereditária.
A mutação e suas conseqüências
Se a alteração na seqüência de aminoácidos na
proteína não afetar o funcionamento da molécula e não prejudicar o
organismo, de modo geral ela passa despercebida, é indiferente.
Outras vezes, a alteração leva a um
favorecimento. Imagine, por exemplo, que uma certa célula do seu
intestino passe a produzir uma enzima chamada celulase, capaz de digerir
a celulose dos vegetais que você come. provavelmente a mutação que
levou a esse erro será vantajosa para você, que poderá eventualmente até
alimentar-se de papel picado.
Muitas vezes, porém, a mutação pode ser prejudicial. Na anemia falciforme,
a substituição do aminoácido ácido glutâmico pelo aminoácido valina, em
uma das cadeias de hemoglobina, conduza a uma alteração na forma da
proteína toda. Essa alteração muda o formato do glóbulo vermelho, que
passa a ser incapaz de transportar oxigênio. Outra conseqüência, grave, é
que hemácias com formato de foice grudam umas nas outras nos capilares
sangüíneos, o que pode provocar obstruções no trajeto para os tecidos.

As mutações são hereditárias
Dependendo da célula em que a mutação ocorre, ela
pode ser transmitida à descendência. Nas suposições que fizemos,
relacionadas ao pigmento da pele e à enzima celulase, evidentemente que
não ocorrerá a transmissão dos genes mutantes para os filhos.
Trata-se de mutações somáticas, ou seja, ocorreram em células não envolvidas na confecção de gametas.
Já a mutação que conduziu à anemia falciforme,
deve ter ocorrido, no passado, em células da linhagem germinativa de
algum antepassado. O gene anômalo, então sugerido, deve ter sido
transportado por um gameta e daí se espalhou pela espécie humana.
As causas das mutações
De maneira geral, as mutações ocorrem como
conseqüência de erro no processo de duplicação do DNA. Acontecem em uma
baixíssima freqüência. Muitas delas, inclusive, são corrigidas por
mecanismos especiais, como, por exemplo, a ação do gene p53 que evita a formação de tumores.
Há, no entanto, certos agentes do ambiente que
podem aumentar a taxa de ocorrência de erros genéticos. Entre esses
agentes mutagênicos podemos citar: substâncias existentes no
fumo, os raios X, a luz ultravioleta, o gás mostarda, ácido nitroso e
algumas corantes existentes nos alimentos. Não é à toa que, em muitos países, é crescente a preocupação com a diminuição da espessura da camada do gás ozônio (O3),
que circunda a atmosfera terrestre. Esse gás atua como filtro de luz
ultravioleta proveniente do Sol. Com a diminuição da sua espessura,
aumenta a incidência desse tipo de radiação, o que pode afetar a pele
das pessoas. Ocorrem lesões no material genético, que podem levar a
certos tipos de câncer de pele.









![[2^24 + 2^30]/65 = x^x](https://blogger.googleusercontent.com/img/b/R29vZ2xl/AVvXsEgDK0elX3csGKExdHvk67b1Q-I81hW4kQDloEJ6Z6g5HhUzYzLg8gybPtqIEwsIHEC2Z3paYRXuRyfaR3sL_EhVE2HAPAHy28-0QClNe8M0ShpLM0ixm-BRK5sqm-kbnJDAUeqHE9sfNEYP3Y7UbCZ87CtsZ0adG5yAc5kwxCo3LnJCDr33oM98C-XUV4d2/w100/Blog%20do%20Professor%20Janildo%20-%20Copia.png)



























0 Comentários